Выполнила студентка 3-го курса ЭГО ЕГФ Гаганова В.В.
Липецкий государственный педагогический университет.
Естественно Географический Факультет.
Кафедра ботаники.
Липецк 2001.
Введение.
Вода – важнейший экологический фактор для всего живого на земле. Для процессов обмена веществ со средой, составляющих основу жизни, необходимо участие воды в качестве растворителя и метаболита. Так у растений вода участвует в реакциях фотосинтеза, минеральные соли поступают в растения из почвы только в виде водных растворов. Вода – главная составная часть тела растений. Даже находясь в анабиозе, растения содержат воду. Особая роль воды наземных растений заключается в постоянном пополнении больших трат ее на испарение в связи с развитием большой фотосинтезирующей поверхности. Вода, обуславливая необходимое тургорное давление, определенным образом участвует и в поддержании формы наземных растений как организмов не имеющих опорного скелета. Также для большой группы растений, живущих в водоемах, морях и океанах, вода является непосредственной средой обитания.
Глава I. Характеристика основных групп растений по отношению к воде.
По приуроченности к местообитаниям с разными условиями увлажнения и выработке соответствующих приспособлений среди наземных растений различают три основных экологических типа: гигрофиты, мезофиты и ксерофиты.
Растения, для которых вода не только необходимый экологический фактор, но непосредственная среда обитания, относятся к водным, называемым гидрофитами.
ГИГРОФИТЫ. Это растения избыточно увлажненных местообитаний с высокой влажностью воздуха и почвы. При довольно большом разнообразии местообитаний, особенностей водного режима и анатомо-морфологических черт всех гигрофитов объединяет отсутствие приспособлений, ограничивающих расход воды, и неспособность выносить даже незначительную ее потерю. Ярко выраженные гигрофиты – травянистые растения и эпифиты влажных тропических лесов, не выносящие сколько–нибудь заметного понижения влажности воздуха. Даже в разгар сезона дождей мелкие эпифитные папоротники на стволах деревьев теряют тургор и засыхают, если на них в течение 2-3 часов падают солнечные лучи. Черты гигрофитов имеют травянистые растения темнохвойных лесов (кислица, майник двулистный, двулепестник альпийский). К гигрофитам можно отнести и виды, растущие на открытых и хорошо освещенных местообитаниях, но в условиях избытка почвенной влаги – близ водоемов, в дельтах рек, в местах выхода грунтовых вод. В наших широтах примером могут служить прибрежные виды: калужница – Caltha palustris, плакун-трава – Lythrum salikaria, а в странах жаркого климата – папирус, болотные пальмы. Из культурных растений сюда можно отнести рис, культивируемый на полях, залитых водой.
КСЕРОФИТЫ. Это растения сухих местообитаний, способные переносить значительный недостаток влаги – почвенную и атмосферную засуху. Они распространены, обильны и разнообразны в областях с жарким и сухим климатом. К этой группе принадлежат виды пустынь, сухих степей, саванн, колючих редколесий, сухих субтропиков. В более гумидных районах ксерофиты участвуют в растительном покрове лишь в наиболее прогреваемых и наименее увлажненных местообитаниях (например, на склонах южной экспозиции).
Неблагоприятный водный режим растений в сухих местообитаниях обусловлен, во-первых, ограниченным поступлением воды при ее недостатке в почве и, во-вторых, увеличением расхода влаги на транспирацию при большой сухости воздуха и высоких температурах. Следовательно, для преодоления недостатка влаги возможны разные пути: увеличение ее поглощения и сокращение расхода, кроме того, способность переносить большие потери воды. Все это используется ксерофитами при адаптации к сухости, но у разных растений в неодинаковой степени, в связи с чем некоторые авторы различают два основных способа преодоления ксерофитами засухи: возможность противостоять иссушению тканей, или активное регулирование водного баланса, и способность выносить сильное иссушение.
В зависимости от структурных черт и способов регулирования водного режима различают несколько разновидностей ксерофитов (по Генкелю П.А.): эуксерофиты, гемиксерофиты, пойкилоксерофиты.
К группе ксерофитов относят и суккуленты – растения с сочными листьями или стеблями. Различают листовые суккуленты (агавы, алоэ) и стеблевые, у которых листья редуцированы, а наземные части представлены мясистыми стеблями(кактусы, некоторые молочаи).
Ксерофиты с наиболее ярко выраженными ксероморфными чертами строения листьев имеют своеобразный внешний облик, за что получили название склерофитов. Облик типичного склерофита легко представить на примере чертополоха – Carduus crispus и пустынных полыней, ковылей, саксаулов.
МЕЗОФИТЫ. Эта группа включает растения, произрастающие в средних условиях увлажнения. Сюда относятся растения лугов, травяного покрова лесов, лиственные древесные и кустарниковые породы из областей умеренно влажного климата, а также большинство культурных растений.
Мезофиты – группа весьма разнообразная не только по видовому составу, но и по различным экологическим оттенкам, обусловленным разным сочетанием факторов в природных местообитаниях. Они связаны переходами с другими экологическими типами растений по отношению к воде, так что четкую границу между ними провести очень трудно. Так, среди луговых мезофитов выделяются виды с повышенным влаголюбием, предпочитающие постоянно сырые или временно заливаемые участки (лисохвост луговой – Alopecurus pratensis, бекмания обыкновенная – Beckmannia eruciformis).
Их объединяют в переходную группу гигромезофитов наряду с некоторыми влаголюбивыми лесными травами, предпочитающими наиболее сырые леса, лесные овраги (недотрога – Impatiens nolitangere). С другой стороны в местообитаниях с переодическим или постоянным (небольшим) недостатком влаги много мезофитов с теми или иными ксероморфными признаками с повышенной физиологической устойчивостью к засухе. Эта группа переходная между мезофитами ксерофитами, - ксеромезофиты. Примером могут служить многие виды северных степей, сухих сосновых боров, песчаных местообитаний: клевер-белоголовка – Trifolium montanum, подмаренник желтый – Galium verum и другие.
Особое место среди мезофитов занимают степные и пустынные весенние эфемеры и эфемероиды. К этой группе принадлежат растения, ранней весной покрывающие степи и пустыни разноцветным цветущим ковром (многолетники – тюльпаны, гусиные луки; однолетники – маки, вероники). Это виды с чрезвычайно краткой вегетацией и длительным периодом покоя, который однолетние эфемеры переживают в виде семян, а многолетние эфемероиды – в виде покоящихся луковиц, клубней, корневищ. Кроме весенних существуют и осенние эфемероиды, произрастающие в районах с климатическим ритмом средиземноморского типа. Сюда относятся виды родов Crocus, Scilla и другие.
По многим особенностям структуры и физиологии близки к ксерофитам растения, которые по тем или иным причинам испытывают недостаток влаги, сопряженный с действием низких температур. Иногда такие виды в качестве особого подразделения включают в группу ксерофитов, иногда выделяют в самостоятельные экологические типы – психрофиты и криофиты.
Психрофиты – растения влажных и холодных почв в холодных местообитаниях высокогорий и северных широт. Несмотря на достаточное увлажнение почвы, они часто испытывают недостаток влаги (или из-за физиологической сухости, вызванной низкими температурами, или в связи с преобладанием в почве недоступной влаги, как, например, на торфянистых почвах). Среди психрофитов есть травянистые растения (например злаки северных лугов: белоус – Nardus strikta; высокогорные кавказские злаки: овсяница пестрая –Festuka varia), высокогорные, болотные и тундровые кустарники и кустарнички, как вечнозеленые (вереск – Calluna vulgaris), так и с опадающей листвой (карликовые ивы – Salix polaris, S. herbacea).К психрофитам относятся и хвойные древесные породы умеренных и северных широт.
Криофиты в экологическом отношении очень близки к психрофитам и связаны с ними переходными формами. Это растения сухих и холодных местообитаний – сухих участков тундр, скал, осыпей. Обычно они рассматриваются и характеризуются вместе с психрофитами, поскольку у них много сходных морфологических и физиологических черт. Но среди криофитов есть и весьма своеобразные формы – это растения-подушки высокогорных холодных пустынь.
ГИДРОФИТЫ. Это водные растения. По образу жизни и строению среди них можно выделить погруженные растения и растения с плавающими листьями. Погруженные растения подразделяют на укореняющиеся в донном грунте и взвешенные в толще воды. Из высших растений к первым принадлежат телорез – Stratiotes aloides, шильник водяной – Subularia aquatika. В эту же группу входят водоросли, прикрепленные к грунту. Из растений, взвешенных в толще воды, можно назвать роголистник погруженный-Ceratophyllum demersum, пузырчатку обыкновенную – Utrikularia vulgaris, а также многочисленные виды планктонных водорослей.
Растения с плавающими листьями используют частично водную, частично воздушную среду. Из них укореняются в грунте кувшинки из рода Nymphaea, кубышки из рода Nuphar, рдесты, орех водяной – Trapa natans.
Многие виды наряду с плавающими на поверхности воды листьями имеют и подводные. Плавают на поверхности воды, не укореняясь, ряски, водокрас.
К настоящим водным растениям очень близко примыкает и обычно вместе с ними рассматривается группа гелофитов или амфибий – земноводных растений. Это виды береговых и прибрежных местообитаний с избыточным или переменным увлажнением. Они могут расти как в воздушной среде, так и частично погруженными в воду, могут выносить и полное временное заливание. Как в природе нет резкой границы между водными и наземными местообитаниями для растений, так и группа гелофитов связана незаметными переходами, с одной стороны, с настоящими гидрофитами, с другой – с наземными гигрофитами и гигромезофитами. Примеры гелофитов – растений прибрежной полосы пресноводных водоемов и рек: стрелолист – Sagittaria sagittifolia, ежеголовка – Sparganium ramosum.
Глава II. Анатомо-морфологические приспособления растений к водному режиму.
Растения, приуроченные к местообитаниям с разными условиями увлажнения, выработали соответствующие приспособления к водному режиму.
Характерные структурные черты гигрофитов – тонкие нежные листовые пластинки с небольшим числом устьиц, не имеющие толстой кутикулы, рыхлое сложение тканей листа с крупными межклетниками, слабое развитие водопроводящей ткани, тонкие слаборазветвленные корни.
Для ксерофитов большое значение имеют разнообразные структурные приспособления к условиям недостатка влаги.
Корневые системы обычно сильно развиты, что помогает растениям увеличить поглощение почвенной влаги. По общей массе корневые системы ксерофитов нередко превышают надземные части, иногда весьма значительно. Так, у многих травянистых и кустарниковых видов среднеазиатских пустынь подземная масса больше надземной в 9-10 раз, а у кесрофитов памирских высокогорных холодных пустынь – в300-400 раз. Корневые системы ксерофитов часто бывают экстенсивного типа, то есть растения имеют длинные корни, распространяющиеся в большом объеме почвы, но сравнительно мало разветвленные. Проникновение таких корней на большую глубину позволяет ксерофитам использовать влагу глубоких почвенных горизонтов, а в отдельных случаях – и грунтовых вод.
У других видов корневые системы интенсивного типа: они охватывают относительно небольшой объем почвы, но благодаря очень густому ветвлению максимально используют почвенную влагу. Корни ряда ксерофильных видов имеют специальные приспособления для запасания влаги. Надземные органы ксерофитов также отличаются своеобразными (так называемыми ксероморфными чертами), которые носят отпечаток трудных условий водоснабжения. У них сильно развита водопроводящая система, что хорошо заметно по густоте сети жилок в листьях, подводящих воду к тканям. Эта черта облегчает ксерофитам пополнение запасов влаги, расходуемой на транспирацию.
Разнообразные структурные приспособления защитного характера, направленные на уменьшение расхода воды, в основном сводятся к следующему:
Общее сокращение транспирирующей поверхности. Многие ксерофиты имеют мелкие, узкие, сильно редуцированные листовые пластинки. В особо засушливых пустынных местообитаниях листья некоторых древесных и кустарниковых пород редуцированы до едва заметных чешуек. У таких видов фотосинтез осуществляют зеленые ветви.
Уменьшение листовой поверхности в наиболее жаркие и сухие периоды вегетационного сезона. Для многих кустарников среднеазиатских, североафриканских и других пустынь, а так же для некоторых видов сухих субтропиков средиземноморья характерен сезонный деформизм листьев: ранней весной при еще благоприятном водном режиме образуются относительно крупные листья, которые летом, при наступлении жары и сухости, сменяются мелкими листьями более ксероморфного строения с меньшей интенсивностью транспирации.
Защита листьев от больших потерь влаги на транспирацию. Она достигается благодаря развитию мощных покровных тканей – толстостенного, иногда многослойного эпидермиса, часто несущего различные выросты и волоски, которые образуют густое “войлочное” опушение поверхности листа. У других видов поверхность покрыта водонепроницаемым слоем толстой кутикулы или воскового налета. Развитие защитных покровов на листьях причина того, что степной травостой имеет тусклые, седоватые оттенки, резко отличающиеся от яркой зелени лугов.
Устьица у ксерофитов обычно защищены от чрезмерной потери влаги, например, расположены в специальных углублениях в ткани листа, иногда снабженных волосками и прочими дополнительными защитными устройствами. У ковылей и других степных злаков существует интересный механизм защиты устьиц в самые жаркие и сухие часы дня: при больших потерях воды крупные тонкостенные водоносные клетки эпидермиса теряют тургор, и лист свертывается в трубку; так устьица оказываются изолированными от окружающего сухого воздуха внутри замкнутой полости, где благодаря транспирации создается повышенная влажность. Во влажную погоду клетки эпидермиса восстанавливают тургор, и листовая пластинка вновь развертывается.
Усиленное развитие механической ткани.
Клетки тканей листьев у ксерофитов отличаются мелкими размерами и весьма плотной упаковкой, то есть малым развитием межклетников, благодаря чему сильно сокращается внутренняя испаряющая поверхность листа. Поскольку ксерофиты обычно обитают на открытых, хорошо освещенных местообитаниях, многие черты ксероморфной структуры листа – это одновременно и черты световой структуры. Так у многих видов листья имеют мощную иногда многорядную палисадную паренхиму, часто расположенную с обеих сторон.
Основные морфолого-анотомические черты мезофитов – средние между чертами гигрофитов и ксерофитов. Мезофиты имеют умеренно развитые корневые системы как экстенсивного, так и интенсивного типа, со всеми переходами между ними. Для листа характерна дифференцировка тканей на более или менее плотную палисадную паренхиму и рыхлую губчатую паренхиму с системой межклетников. Сеть жилок сравнительно негустая. Покровные ткани могут иметь отдельные ксероморфные черты, но не столь ярко выраженные, как у ксерофитов.
Психрофиты имеют ярко выраженную ксероморфную структуру листа. Так, психрофильные злаки узколистны, имеют хорошо развитую проводящую и механическую ткани; некоторые из них способны к свертыванию листовой пластинки в трубку, напоминая степные ксерофиты. Вечнозеленые кустарнички имеют плотные кожистые листья, иногда с весьма мощной кутикулой, плотной палисадной паренхимой. Нижняя сторона, несущая устьица, часто опушена или покрыта восковым налетом. Вместе с тем в структуре листа ясно заметны и некоторые гигроморфные черты, главным образом, крупные размеры клеток и развитие больших межклетников в губчатой ткани.
Анатомо-морфологические черты гидрофитов существенно отличают их от наземных растений. Заметна редукция механических тканей. Столь же сильно редуцированна и проводящая система. Если у сухопутных мезофитов длина жилок на 1 кв.см. листа составляет около100 мм и более, а у ксерофитов доходит до300, то у водных и прибрежных растений она в несколько раз меньше. Вот несколько примеров (по Гесснеру Ф., 1959):
У некоторых погруженных растений, не прикрепленных к грунту, корни полностью редуцированны, другие корни сохранили, но отдельно плавающие части растений могут обходиться и без них. Корни укрепляющихся гидрофитов слабоветвистые, без корневых волосков. Вместе с тем ряд видов имеет толстые и прочные корневища, которые играют роль якоря, хранилища запасных веществ и органа вегетативного размножения.
Листья погруженных гидрофитов очень тонки и нежны, имеют упрощенное строение мезофилла без заметной дифференциации на палисадную и губчатую паренхиму. Подводные листья без устьиц. В отдельных местах находятся группы клеток эпидермиса с утонченными стенками. Считается, что они играют большую роль в поглощении воды и растворенных минеральных солей.
У растений лишь частично погруженных в воду, хорошо выражена гетерофиллия - различие строения надводных и подводных листьев на одной и той же особи. Первые имеют черты, обычные для листьев наземных растений, вторые – очень тонкие или рассеченные листовые пластинки. Гетерофиллия отмечена у водного лютика – Ranunculus diversifolius, кувшинок и кубышек, стрелолиста и других видов. Интересный пример – поручейник, на стебле которого можно видеть несколько форм листьев, представляющих все переходы от типично наземных до водных.
Глава III.Физиологические адаптации растений, приуроченных к местообитаниям разной увлажненности.
Наряду с морфологическими особенностями у растений, приуроченных к местам с разными условиями увлажненности, выработались и физиологические.
Способность гигрофитов к регуляции водного режима ограничена: устьица большей частью широко открыты, так что транспирация мало отличается от физического испарения. Благодаря беспрепятственному потоку воды и отсутствию защитных приспособлений интенсивность транспирации очень высока: у световых гигрофитов в дневное время листья могут терять за час количество воды, в 4-5 раз превышающее массу листа. Высокая оводненность тканей гигрофитов поддерживается в основном за счет постоянного притока влаги из окружающей среды.
Другие характерные физиологические черты гигрофитов, обусловленные легкой доступностью влаги, - низкое осмотическое давление клеточного сока, незначительная водоудерживающая способность, приводящая к быстрой потере запасов воды.особенно показательны для гигрофитов небольшие величины сублетального водного дефицита: так, для кислицы и майника потеря 15%-20% запаса воды уже необратима и ведет к гибели. В некоторых случаях у растений сильно увлажненных местообитаний возникает необходимость удаления избытка влаги. Обычно, это бывает, когда почва хорошо прогрета и корни активно всасывают воду, а транспирация отсутствует (например, утром или при тумане, когда влажность воздуха 100%). Избыточная влага удаляется путем гуттации – выделение воды через специальные выделительные клетки, расположенные по краю или на острие листа.
Ксерофиты обладают рядом разнообразных физиологических адаптаций, позволяющих им успешно выдерживать недостаток влаги.
У ксерофитов обычно повышено осмотическое давление клеточного сока, позволяющее всасывать воду даже при больших водоотнимающих силах почвы, то есть использовать не только легкодоступную, но и труднодоступную почвенную влагу. Оно измеряется тысячами кПа, а у некоторых пустынных кустарников зарегистрированы цифры, достигающие 10000-30000 кПа.
С давних пор пристальное внимание привлекала проблема расхода воды ксерофитами на транспирацию. Казалось бы, многочисленные анатомические приспособления, достаточно надежно защищающие наземные части ксерофитов от сильного испарения, должны способствовать значительному снижению транспирации. Однако выяснилось, что в действительности это не так. При достаточном водоснабжении большинство ксерофитов имеют довольно высокую транспирацию, но при наступлении засушливых условий, они сильно сокращают ее. При этом играет роль и закрывание устьиц, и сильное обезвоживание листа при начинающемся подвядании. Несомненно, анатомо-морфологические приспособления имеют определенное значение, но основную роль в засухоустойчивости ксерофитов в настоящее время отводят физиологическим механизмам.
К числу этих механизмов принадлежит высокая водоудерживающая способность тканей и клеток, обусловленная рядом физиологических и биохимических особенностей.
Большое значение для выживания ксерофитов при резком недостатке влаги имеет их способность переносить глубокое обезвоживание тканей без потери жизнеспособности и способности восстановления нормального содержания воды в растении при возобновлении благоприятных условий. Ксерофиты способны потерять до 75% всего водного запаса и, тем не менее, остаться живыми. Ярким примером в этом отношении служат пустынные растения, которые летом высыхают до состояния, близкого к воздушно-сухому, и впадают в анабиоз, но после дождей возобновляют рост и развитие.
Еще одна система адаптаций, обеспечивающих выживание ксерофитов в аридных условиях, - выработка сезонных ритмов, дающих возможность растениям использовать для вегетации наиболее благоприятные периоды года и резко сократить жизнедеятельность во время засухи. Так, в областях со средиземноморским климатом с резко выраженным летним сухим периодом многие ксерофильные виды имеют “двухтактный” ритм сезонного развития: весенняя вегетация сменяется летним покоем, во время которого растения сбрасывают листву и снижают интенсивность физиологических процессов; в период осенних дождей вегетация возобновляется, и затем уже следует зимний покой. Сходное явление наблюдается и у растений сухих степей в середине и конце лета: потеря части листовой поверхности, приостановка развития, сильное обезвоживание тканей и т.д. Такое состояние, получившее название полупокоя, длится вплоть до осенних дождей, после которых у степных ксерофитов начинают отрастать листья.
Физиологические адаптации суккулентов столь своеобразны, что их необходимо рассмотреть отдельно.
Основной способ преодоления засушливых условий у суккулентов – накопление больших запасов воды в тканях и крайне экономное ее расходование. В условиях жаркого и сухого климата весь водный запас мог бы быть быстро растрачен, но растения имеют защитные приспособления, направленные к сокращению транспирации. Одно из них – своеобразная форма надземных частей суккулентов. В дополнение к этому у многих суккулентов поверхность защищена восковым налетом опушением, хотя есть и суккуленты с тонким не защищенным эпидермисом. Устьица очень немногочисленны, часто погружены в ткань листа или стебля. Днем устьица обычно закрыты, и потеря воды идет в основном через покровные ткани.
Транспирация у суккулентов чрезвычайно мала. Ее трудно уловить за короткий период и приходится определять расход воды не за час, а за сутки или за неделю. Водоудерживающая способность тканей суккулентов значительно выше, чем у других растений экологических групп, благодаря содержанию в клетках гидрофильных веществ. Поэтому и без доступа влаги суккуленты расходуют водный запас очень медленно и долго сохраняют жизнеспособность даже в гербарии.
Ограничения, обусловленные особенностями водного режима суккулентов, создают и другие трудности для жизни этих растений в аридных условиях. Слабая транспирация сводит к минимуму возможность терморегуляции, с чем связано сильное нагревание массивных надземных органов суккулентов. Затруднения создаются и для фотосинтеза, поскольку днем устьица обычно закрыты, а открываются ночью, следовательно, доступ углекислоты и света не совпадают во времени. Поэтому у суккулентов выработался особый путь фотосинтеза, при котором в качестве источника углекислоты, частично используются продукты дыхания. Иными словами, в крайних условиях растения частично используют принцип замкнутой системы с реутилизацией отходов метаболизма.
В силу всех этих ограничений интенсивность фотосинтеза суккулентов невелика, рост и накопление массы идут очень медленно, вследствие чего они не отличаются высокой биологической продуктивностью и не образуют сомкнутых растительных сообществ.
Физиологические показатели водного режима мезофитов подтверждают их промежуточную позицию: для них характерны умеренные величины осмотического давления, содержания воды в листьях, предельного водного дефицита. Что касается транспирации, то ее величина в большей степени зависит от условий освещенности и других элементов микро климата.
Один и тот же мезофильный вид, попадая в разные по водоснабжению условия, обнаруживает известную пластичность, приобретая в сухих условиях более ксероморфные, а во влажных более гигроморфные черты.
Пластичность листьев проявляется не только в разных местообитаниях, но даже у одной и той же особи. Например, у деревьев на опушке леса листья на стороне, обращенной в сторону леса, имеют более мезофильный и теневой характер по сравнению с несколько ксероморфными листьями внешней стороны дерева (см. табл. ниже). Листья разных высотных ярусов одних и тех же растений находятся в неодинаковых условиях водоснабжения, так как поступление воды в верхние части связано с преодолением большого сопротивления. К тому же у деревьев верхние листья обычно находятся в условиях иного микроклимата.
Различие анатомо-физиологических показателей листьев на разных сторонах кроны дерева, растущего на опушке леса.
I – сторона, обращенная к лесу, II – сторона, обращенная к поляне
Древесная порода
Площадь листа, см 2
Число устьиц на
Содержание воды, %
Содержание хлорофилла, мг/г
Средняя интенсивность фотосинтеза, мг СО 2 /дм 2 ·ч
Толщина листа, мкм
Дуб – Quercus robur
I
42
45
61
3.0
1.9
97
II
18
125
54
2.4
2.3
181
Липа – Tilia cordata
I
38
38
71
3.6
1.6
93
II
24
45
62
2.0
1.1
106
Водная среда существенно отличается от воздушной, поэтому у водных растений существует ряд своеобразных физиологических адаптивных черт. Интенсивность света в воде сильно ослаблена, поскольку часть падающей радиации отражается от поверхности воды, другая – поглощается ее толщей. В связи с ослаблением света фотосинтез у погруженных растений сильно снижается с увеличением глубины. Считают, сто выживанию глубоководного фитопланктона в зонах, где освещенность ниже точки компенсации, способствуют его периодические вертикальные перемещения в верхние зоны, где идет интенсивный фотосинтез и пополнение запасов органических веществ.
В воде кроме недостатка света растения могут испытывать и другое затруднение, существенное для фотосинтеза, - недостаток доступной СО 2 . Углекислота поступает в воду в результате растворения СО 2 , содержащегося в воздухе, дыхания водных организмов, разложения органических остатков и высвобождения из карбонатов. При интенсивном фотосинтезе растений идет усиленное потребление СО 2 , в связи с чем легко возникает ее дефицит. На увеличение содержания СО 2 в воде гидрофиты реагируют заметным повышением фотосинтеза.
У погруженных растений транспирации нет, значит, нет и “верхнего двигателя”, поддерживающего ток воды в растении. Однако этот ток, доставляющий к тканям питательные вещества, существует, при чем с явной суточной периодичностью: днем больше, ночью отсутствует. Активная роль в его поддержании принадлежит корневому давлению и деятельности специальных клеток, выделяющих воду, - водяных устьиц.
Плавающие или торчащие над водой листья обычно имеют сильную транспирацию, хотя и расположены в слое воздуха, который непосредственно граничит с водой и имеет повышенную влажность. Устьица широко открыты и закрываются полностью только в ночное время.
Столь же велика транспирация у прибрежных растений, при чем у них значительное количество воды расходуется не только листьями, но и стеблями.
Осмотическое давление у водных и прибрежных растений очень низкое, так как им не приходится преодолевать водоудерживающую силу почвы при поглощении воды.
Примеры осмотического давления (в кПа) у водных растений и гелофитов (по Гесснеру Ф.,1959)
Список литературы
Ботаника с основами экологии: Учеб. Пособие для студентов пед. ин-тов /Л.В. Кудряшов и др. – М.: Просвещение,1979
Горышина Т.К. Экология растений. – М.: Высшая школа,1979
Гусев Н.А. Некоторые закономерности водного режима растений. – М.: Изд-во АН СССР, 1959.
Двораковский М.С. Экология растений. – М.: Просвещение 1964
Крафтс А. Вода и ее значение в жизни растений. – М.: Иност. лит., 1951
Культиасов И.М. Экология растений. - М.: Изд-во московского ун-та, 1982
Мак-Дуголл В.Б. Экология растений. – М.: Учпедгиз, 1935.
Пономарева И.Н. Экология растений с основами биогеоценологии. – М.: Просвещение, 1978
Рычин Ю.В. Флора гигрофитов. Определитель. – М.: Сов. наука, 1948
Шенников А.П. Экология растений. – М.: Сов. наука, 1950
Часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
У засухоустойчивых растений эти показатели будут выше. В селекционной работе используют такой показатель, как содержание статолитного крахмала в корневом чехлике. Генетически обусловленным признаком засухоустойчивости растений является способность их вегетативных органов (особенно листьев) накапливать во время засухи пролин. При этом концентрация пролина увеличивается в 10-100 раз. В пролине...
И гибридов тех или иных сельскохозяйственных культур может различаться и по качеству урожая – содержанию белка, клейковины, крахмала, сахара, жира и т.д. Однако при агроэкологической оценке сельскохозяйственных культур в конкретных почвенно-климатических условиях необходимо использовать региональные справочные материалы, что позволит свести к минимуму ошибки в оптимизации структуры посевных...
Опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется. Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация. Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, ...
Министерство общего и
профессионального образования Р.Ф.
Липецкий государственный педагогический
университет.
Естественно Географический Факультет.
Кафедра ботаники.
Курсовая работа
на тему:
«Приспособление растений к водному режиму»
Работу
выполнила студентка
3-го курса ЭГО ЕГФ:
Гаганова В.В.
Работу приняла:
Соловьева Н.Ю.
Липецк
2001.
Введение.
…………………………………………………………….3
Глава
I
.
Характеристика основных групп растений по отношению к воде. …………….………………………………………...4
Глава
II
.
Анатомо-морфологические приспособления
растений к водному режиму. ………………….……………………..10
Глава
III
.
Физиологические
адаптации растений, приуроченных к местообитаниям разной увлажненности.
………..16
Список
литературы.
………………………………………………24
Введение.
Вода – важнейший экологический фактор для всего живого на земле. Для
процессов обмена веществ со средой, составляющих основу жизни, необходимо
участие воды в качестве растворителя и метаболита. Так у растений вода
участвует в реакциях фотосинтеза, минеральные соли поступают в растения из
почвы только в виде водных растворов. Вода – главная составная часть тела
растений. Даже находясь в анабиозе, растения содержат воду. Особая роль воды
наземных растений заключается в постоянном пополнении больших трат ее на
испарение в связи с развитием большой фотосинтезирующей поверхности. Вода,
обуславливая необходимое тургорное давление, определенным образом участвует и в
поддержании формы наземных растений как организмов не имеющих опорного скелета.
Также для большой группы растений, живущих в водоемах, морях и океанах, вода
является непосредственной средой обитания.
Глава
I. Характеристика
основных групп растений по отношению к воде.
По приуроченности к местообитаниям с разными условиями увлажнения и
выработке соответствующих приспособлений среди наземных растений различают три
основных экологических типа: гигрофиты, мезофиты и ксерофиты.
Растения, для которых вода не только необходимый экологический фактор,
но непосредственная среда обитания, относятся к водным, называемым гидрофитами.
ГИГРОФИТЫ.
Это растения избыточно увлажненных
местообитаний с высокой влажностью воздуха и почвы. При довольно большом
разнообразии местообитаний, особенностей водного режима и
анатомо-морфологических черт всех гигрофитов объединяет отсутствие
приспособлений, ограничивающих расход воды, и неспособность выносить даже
незначительную ее потерю. Ярко выраженные гигрофиты – травянистые растения и
эпифиты влажных тропических лесов, не выносящие сколько–нибудь заметного
понижения влажности воздуха. Даже в разгар сезона дождей мелкие эпифитные
папоротники на стволах деревьев теряют тургор и засыхают, если на них в течение
2-3 часов падают солнечные лучи. Черты гигрофитов имеют травянистые растения
темнохвойных лесов (кислица, майник двулистный, двулепестник альпийский). К
гигрофитам можно отнести и виды, растущие на открытых и хорошо освещенных
местообитаниях, но в условиях избытка почвенной влаги – близ водоемов, в
дельтах рек, в местах выхода грунтовых вод. В наших широтах примером могут
служить прибрежные виды: калужница – Caltha palustris,
плакун-трава – Lythrum salikaria, а в странах жаркого
климата – папирус, болотные пальмы. Из культурных растений сюда можно отнести
рис, культивируемый на полях, залитых водой.
КСЕРОФИТЫ.
Это растения сухих местообитаний,
способные переносить значительный недостаток влаги – почвенную и атмосферную
засуху. Они распространены, обильны и разнообразны в областях с жарким и сухим
климатом. К этой группе принадлежат виды пустынь, сухих степей, саванн, колючих
редколесий, сухих субтропиков. В более гумидных районах ксерофиты участвуют в
растительном покрове лишь в наиболее прогреваемых и наименее увлажненных
местообитаниях (например, на склонах южной экспозиции).
Неблагоприятный водный режим растений в сухих местообитаниях
обусловлен, во-первых, ограниченным поступлением воды при ее недостатке в почве
и, во-вторых, увеличением расхода влаги на транспирацию при большой сухости
воздуха и высоких температурах. Следовательно, для преодоления недостатка влаги
возможны разные пути: увеличение ее поглощения и сокращение расхода, кроме
того, способность переносить большие потери воды. Все это используется
ксерофитами при адаптации к сухости, но у разных растений в неодинаковой
степени, в связи с чем некоторые авторы различают два основных способа
преодоления ксерофитами засухи: возможность противостоять иссушению тканей, или
активное регулирование водного баланса, и способность выносить сильное
иссушение.
В зависимости от структурных черт и способов регулирования водного
режима различают несколько разновидностей ксерофитов (по Генкелю П.А.):
эуксерофиты, гемиксерофиты, пойкилоксерофиты.
К группе ксерофитов относят и суккуленты – растения с сочными листьями
или стеблями. Различают листовые суккуленты (агавы, алоэ) и стеблевые, у
которых листья редуцированы, а наземные части представлены мясистыми
стеблями(кактусы, некоторые молочаи).
Ксерофиты с наиболее ярко выраженными ксероморфными чертами строения
листьев имеют своеобразный внешний облик, за что получили название склерофитов.
Облик типичного склерофита легко представить на примере чертополоха – Carduus
crispus и пустынных полыней, ковылей, саксаулов.
МЕЗОФИТЫ.
Эта группа включает растения,
произрастающие в средних условиях увлажнения. Сюда относятся растения лугов,
травяного покрова лесов, лиственные древесные и кустарниковые породы из
областей умеренно влажного климата, а также большинство культурных растений.
Мезофиты – группа весьма разнообразная не только по видовому составу,
но и по различным экологическим оттенкам, обусловленным разным сочетанием
факторов в природных местообитаниях. Они связаны переходами с другими
экологическими типами растений по отношению к воде, так что четкую границу
между ними провести очень трудно. Так, среди луговых мезофитов выделяются виды
с повышенным влаголюбием, предпочитающие постоянно сырые или временно
заливаемые участки (лисохвост луговой – Alopecurus pratensis, бекмания обыкновенная – Beckmannia eruciformis).
Их
объединяют в переходную группу гигромезофитов наряду с некоторыми влаголюбивыми
лесными травами, предпочитающими наиболее сырые леса, лесные овраги (недотрога
– Impatiens nolitangere). С другой стороны в
местообитаниях с переодическим или постоянным (небольшим) недостатком влаги
много мезофитов с теми или иными ксероморфными признаками с повышенной
физиологической устойчивостью к засухе. Эта группа переходная между мезофитами
ксерофитами, - ксеромезофиты. Примером могут служить многие виды северных
степей, сухих сосновых боров, песчаных местообитаний: клевер-белоголовка – Trifolium
montanum, подмаренник желтый – Galium verum
и другие.
Особое
место среди мезофитов занимают степные и пустынные весенние эфемеры и
эфемероиды. К этой группе принадлежат растения, ранней весной покрывающие степи
и пустыни разноцветным цветущим ковром (многолетники – тюльпаны, гусиные луки;
однолетники – маки, вероники). Это виды с чрезвычайно краткой вегетацией и
длительным периодом покоя, который однолетние эфемеры переживают в виде семян,
а многолетние эфемероиды – в виде покоящихся луковиц, клубней, корневищ. Кроме
весенних существуют и осенние эфемероиды, произрастающие в районах с
климатическим ритмом средиземноморского типа. Сюда относятся виды родов Crocus, Scilla и другие.
По многим
особенностям структуры и физиологии близки к ксерофитам растения, которые по
тем или иным причинам испытывают недостаток влаги, сопряженный с действием
низких температур. Иногда такие виды в качестве особого подразделения включают
в группу ксерофитов, иногда выделяют в самостоятельные экологические типы –
психрофиты и криофиты.
Психрофиты
– растения влажных и холодных почв в холодных местообитаниях высокогорий и
северных широт. Несмотря на достаточное увлажнение почвы, они часто испытывают
недостаток влаги (или из-за физиологической сухости, вызванной низкими
температурами, или в связи с преобладанием в почве недоступной влаги, как,
например, на торфянистых почвах). Среди психрофитов есть травянистые растения
(например злаки северных лугов: белоус – Nardus strikta;
высокогорные кавказские злаки: овсяница пестрая –Festuka varia), высокогорные, болотные и тундровые кустарники и кустарнички, как
вечнозеленые (вереск – Calluna vulgaris), так и с опадающей
листвой (карликовые ивы – Salix polaris, S. herbacea).К психрофитам относятся и хвойные
древесные породы умеренных и северных широт.
Криофиты в
экологическом отношении очень близки к психрофитам и связаны с ними переходными
формами. Это растения сухих и холодных местообитаний – сухих участков тундр,
скал, осыпей. Обычно они рассматриваются и характеризуются вместе с
психрофитами, поскольку у них много сходных морфологических и физиологических
черт. Но среди криофитов есть и весьма своеобразные формы – это
растения-подушки высокогорных холодных пустынь.
ГИДРОФИТЫ.
Это водные растения. По образу жизни и строению среди них можно
выделить погруженные растения и растения с плавающими листьями. Погруженные
растения подразделяют на укореняющиеся в донном грунте и взвешенные в толще
воды. Из высших растений к первым принадлежат телорез – Stratiotes aloides, шильник водяной – Subularia aquatika. В эту же
группу входят водоросли, прикрепленные к грунту. Из растений, взвешенных в
толще воды, можно назвать роголистник погруженный-Ceratophyllum demersum, пузырчатку обыкновенную – Utrikularia vulgaris,
а также многочисленные виды планктонных водорослей.
Растения с
плавающими листьями используют частично водную, частично воздушную среду. Из
них укореняются в грунте кувшинки из рода Nymphaea,
кубышки из рода Nuphar, рдесты, орех водяной – Trapa
natans.
Многие
виды наряду с плавающими на поверхности воды листьями имеют и подводные.
Плавают на поверхности воды, не укореняясь, ряски, водокрас.
К
настоящим водным растениям очень близко примыкает и обычно вместе с ними
рассматривается группа гелофитов или амфибий – земноводных растений. Это виды
береговых и прибрежных местообитаний с избыточным или переменным увлажнением.
Они могут расти как в воздушной среде, так и частично погруженными в воду,
могут выносить и полное временное заливание. Как в природе нет резкой границы
между водными и наземными местообитаниями для растений, так и группа гелофитов
связана незаметными переходами, с одной стороны, с настоящими гидрофитами, с другой
– с наземными гигрофитами и гигромезофитами. Примеры гелофитов – растений
прибрежной полосы пресноводных водоемов и рек: стрелолист – Sagittaria sagittifolia, ежеголовка – Sparganium ramosum.
ГЛАВА
II.
Анатомо-морфологические приспособления растений к водному режиму.
Растения, приуроченные к местообитаниям с разными условиями увлажнения,
выработали соответствующие приспособления к водному режиму.
Характерные структурные черты гигрофитов
– тонкие
нежные листовые пластинки с небольшим числом устьиц, не имеющие толстой
кутикулы, рыхлое сложение тканей листа с крупными межклетниками, слабое
развитие водопроводящей ткани, тонкие слаборазветвленные корни.
Для ксерофитов
большое значение имеют
разнообразные структурные приспособления к условиям недостатка влаги.
Корневые системы обычно сильно развиты, что помогает растениям
увеличить поглощение почвенной влаги. По общей массе корневые системы
ксерофитов нередко превышают надземные части, иногда весьма значительно. Так, у
многих травянистых и кустарниковых видов среднеазиатских пустынь подземная
масса больше надземной в 9-10 раз, а у кесрофитов памирских высокогорных
холодных пустынь – в300-400 раз. Корневые системы ксерофитов часто бывают
экстенсивного типа, то есть растения имеют длинные корни, распространяющиеся в
большом объеме почвы, но сравнительно мало разветвленные. Проникновение таких
корней на большую глубину позволяет ксерофитам использовать влагу глубоких
почвенных горизонтов, а в отдельных случаях – и грунтовых вод.
У других видов корневые системы интенсивного типа: они охватывают
относительно небольшой объем почвы, но благодаря очень густому ветвлению
максимально используют почвенную влагу. Корни ряда ксерофильных видов имеют
специальные приспособления для запасания влаги. Надземные органы ксерофитов
также отличаются своеобразными (так называемыми ксероморфными чертами), которые
носят отпечаток трудных условий водоснабжения. У них сильно развита
водопроводящая система, что хорошо заметно по густоте сети жилок в листьях,
подводящих воду к тканям. Эта черта облегчает ксерофитам пополнение запасов
влаги, расходуемой на транспирацию.
Разнообразные структурные приспособления защитного характера,
направленные на уменьшение расхода воды, в основном сводятся к следующему:
1.
Общее сокращение транспирирующей
поверхности. Многие ксерофиты имеют мелкие, узкие, сильно редуцированные
листовые пластинки. В особо засушливых пустынных местообитаниях листья
некоторых древесных и кустарниковых пород редуцированы до едва заметных чешуек.
У таких видов фотосинтез осуществляют зеленые ветви.
2.
Уменьшение листовой поверхности в наиболее
жаркие и сухие периоды вегетационного сезона. Для многих кустарников
среднеазиатских, североафриканских и других пустынь, а так же для некоторых
видов сухих субтропиков средиземноморья характерен сезонный деформизм листьев:
ранней весной при еще благоприятном водном режиме образуются относительно
крупные листья, которые летом, при наступлении жары и сухости, сменяются
мелкими листьями более ксероморфного строения с меньшей интенсивностью
транспирации.
3.
Защита листьев от больших потерь влаги на
транспирацию. Она достигается благодаря развитию мощных покровных тканей –
толстостенного, иногда многослойного эпидермиса, часто несущего различные
выросты и волоски, которые образуют густое “войлочное” опушение поверхности
листа. У других видов поверхность покрыта водонепроницаемым слоем толстой
кутикулы или воскового налета. Развитие защитных покровов на листьях причина
того, что степной травостой имеет тусклые, седоватые оттенки, резко отличающиеся
от яркой зелени лугов.
Устьица у ксерофитов обычно защищены
от чрезмерной потери влаги, например, расположены в специальных
углублениях в ткани листа, иногда снабженных волосками и прочими
дополнительными защитными устройствами. У ковылей и других степных злаков
существует интересный механизм защиты устьиц в самые жаркие и сухие часы дня:
при больших потерях воды крупные тонкостенные водоносные клетки эпидермиса
теряют тургор, и лист свертывается в трубку; так устьица оказываются
изолированными от окружающего сухого воздуха внутри замкнутой полости, где
благодаря транспирации создается повышенная влажность. Во влажную погоду клетки
эпидермиса восстанавливают тургор, и листовая пластинка вновь развертывается.
4.
Усиленное развитие механической ткани.
Клетки тканей листьев у ксерофитов отличаются мелкими размерами и
весьма плотной упаковкой, то есть малым развитием межклетников, благодаря чему
сильно сокращается внутренняя испаряющая поверхность листа. Поскольку ксерофиты
обычно обитают на открытых, хорошо освещенных местообитаниях, многие черты
ксероморфной структуры листа – это одновременно и черты световой структуры. Так
у многих видов листья имеют мощную иногда многорядную палисадную паренхиму,
часто расположенную с обеих сторон.
Основные
морфолого-анотомические черты мезофитов
– средние между чертами
гигрофитов и ксерофитов. Мезофиты имеют умеренно развитые корневые системы как
экстенсивного, так и интенсивного типа, со всеми переходами между ними. Для
листа характерна дифференцировка тканей на более или менее плотную палисадную
паренхиму и рыхлую губчатую паренхиму с системой межклетников. Сеть жилок
сравнительно негустая. Покровные ткани могут иметь отдельные ксероморфные
черты, но не столь ярко выраженные, как у ксерофитов.
Психрофиты
имеют ярко выраженную ксероморфную структуру листа. Так, психрофильные злаки
узколистны, имеют хорошо развитую проводящую и механическую ткани; некоторые
из них способны к свертыванию листовой пластинки в трубку, напоминая степные
ксерофиты. Вечнозеленые кустарнички имеют плотные кожистые листья, иногда с
весьма мощной кутикулой, плотной палисадной паренхимой. Нижняя сторона, несущая
устьица, часто опушена или покрыта восковым налетом. Вместе с тем в структуре
листа ясно заметны и некоторые гигроморфные черты, главным образом, крупные
размеры клеток и развитие больших межклетников в губчатой ткани.
Анатомо-морфологические
черты гидрофитов
существенно отличают их от наземных растений.
Заметна редукция механических тканей. Столь же сильно редуцированна и
проводящая система. Если у сухопутных мезофитов длина жилок на 1 кв.см. листа
составляет около100 мм и более, а у ксерофитов доходит до300, то у водных и
прибрежных растений она в несколько раз меньше. Вот несколько примеров (по
Гесснеру Ф., 1959).
1.1 Основные приспособления высших растений к водной среде.
1.2. Приспособление растения к водной среде.
Глава II Исследование флоры и фауны пруда.
2.1. Эколог - биологические группы водных растений.
2.2. Эколого-биологический анализ.
2.4 Изучение фауны пруда.
Обсуждение исследуемой работы.
Выводы
Список использованной литературы
Приложение
Введение.
Пруд- сложная природная система то есть комплекс совместно живущих и связанных друг с другом видов .В пруде совместно живущие виды оказываются приспособленными к определенному комплексу абиотических условий и поддерживают свое существо через связи друг с другом. Все виды приспособились к совместному обитанию в ходе длительной эволюции. Отдельные виды связаны между собой прямыми или косвенными пищевыми отношениями и имеют сложную внутреннюю структуру, созданной средой обитания друг для друга и взаимно ограничивают численность. В пруду как в любом биоценозе происходят процессы борьбы за существование и естественного отбора. Водные растения являются первоисточником различных биологических ресурсов водоемов. От них зависит состояние водоемов, разнообразие и обилие населяющих их животных, так как для многих из них растения служат источником питания, для других местом прикрепления икры, для третьих являются убежищем, местом гнездования птиц. В современных условиях водные растения фактически остаются невостребованными вследствие их слабой изученности и приоритетности других. Все вместе это обуславливает актуальность более интенсивного изучения водных растений: их биологии, структурных и функциональных особенностей, экологии их сообществ, что представляет не только теоретический интерес, но и имеет важное практическое значение.
Поскольку видовой состав обитателей прудов нашей местности мало изучен я поставила перед собой цель:
Изучить видовой состав флоры и фауны «Рыбного пруд».
Были поставлены следующие задачи:
1) провести опись флоры и фауны пруда на месте
2) провести систематический и эколого-биологический анализы флоры и фауны.
Глава 1.
1.КРАТКАЯ ХАРАКТЕРИСТИКА рыбного пруда.
Наши исследования проводились июне –июле 2008 года. Объектом нашего исследования был рыбный пруд расположенный в 2-х км от с.Шимкусы. Протяженность пруда составляет -378 м, наибольшая ширина- 5м. Наибольшая глубина достигала 1.5 м. Дно пруда илистое, слой ила местами доходит, по нашим данным, до 90 см. Вода имеет зеленоватый цвет.
Растения и животные, населяющие пруд, распределяются в нем неравномерно. Каждый вид обитает в тех условиях , к которым приспособлен. Наиболее разнообразные и благоприятные для жизни условия создаются в прибрежной зоне. пруда Здесь вода теплее, так как прогревается солнечными лучами. Она достаточна, насыщена кислородом. Обилие света, проникающего до дна, обеспечивает развитие многих высших растений (осока водяная, рдест плавающий, камыш озёрный, рогоз широколистный, и.т.д.). Многочисленны и мелкие водоросли. В прибрежной зоне живут и большинство животных. Одни приспособлены к жизни на водных растениях, другие активно плавают в толщине воды (рыбы, хищные жуки –плавунцы и водяные клопы). Многие водятся на дне (личинки некоторых насекомых- ручейников, стрекоз, поденок, ряд червей и т. п.) . Даже поверхностная плёнка воды служит местом обитания специально приспособленных к ней видов. В тихих заводях можно видеть бегающих по поверхности воды хищных клопов-водомерок и быстро плавающих кругами жуков – вертячек.
1.1Основные приспособления высших растений к водной среде обитания.
Водная среда одновременно влияет и на форму растительных органов и на их жизненные отправления; одно и другое находятся в тесной зависимости.
Первое, что сказывается на жизни и строении водных растений, ¾ это сама водная среда; вода со всех сторон окружает растения, погруженные в нее, и служит основанием, так сказать пьедесталом, для растений, плавающих на ее поверхности.
Вместе с таким быстрым ростом происходит угнетение полового процесса, и размножение сводится исключительно вегетативному это и понятно: если рост и вегетативное размножение идут так успешно, то половое размножение уже теряет свое значение; например: ряски.
Жизнь в водной среде обуславливает иные условия; принятие питательных веществ и перенос их по органам растения проходит совершенно иначе, чем у наземных растений.
Отметим некоторые основные приспособления высших растений к водной среде обитания:
1. Сравнительно низкая температура воды вызывает угнетение полового процесса, преобладает вегетативное размножение.
Замечательные приспособления видим мы у водных растений по отношению к температурным условиям. В общем, жизнь водных растений подчиняется несколько иному температурному режиму по сравнению с наземными: здесь весной и летом холоднее, чем на воздухе, зато зимой теплее подо льдом, в не замёрзшей воде. Это позволяет некоторым из водных растений зимовать без всяких приспособлений, опускаясь на дно водоёма, как ряска, или же продолжая своё существование на дне, как болотники некоторые рдесты. Реже перезимовывает растение в виде клубня: таковы стрелолист и рдест гребенчатый. Чаще всего растение перезимовывает в виде особых зимующих почек, которые представляют собою укороченные побег, с сильно скученными укороченными листьями. Такие побеги, опускаясь на дно, весной дают начало новым особям. Мы видим их у пузырчатки, водокраса, ряски и ряда другие.
2. Усиленный рост по сравнению с наземными растениями , так как в течение сравнительно короткого вегетационного периода растение должно развиться, дать семена или зимующие почки и запасти питательные вещества в подземных органах на зимний период.
3. Отсутствие потребности в прохождении воды от корневой системы к листовым органам вызвало у водных растений недоразвитие древесины в сосудистых пучках, которое в одних случаях (например у роголистника) даже и не закладывается вовсе, а в других случаях недоразвивается.
Недоразвитие или отсутствие древесины в сосудистых пучках, связанное с тем, что растение, поддерживаемое водой, не нуждается в такой мере в опорных элементах , как наземное. Механические элементы у ряда видов, обеспечивающие гибкость стеблей и листьев при сильном течении или волнении, расположены, в отличие от растений суши ближе к центру стебля и по центральной оси листа. Развитие системы воздухоносных полостей (аэренхима) способствует улучшению газообмена и поддерживанию растения в плавающем состоянии.
4. Редукция корневой системы или изменение ее функции. Так, корень или корневидные образования рясок - прежде всего орган равновесия, способствующим плаванию растений на поверхности водоема. Хорошо развитые корневые системы нимфейных служат как для прикрепления их к грунту, так и для запасания питательных веществ.
5. Гетерофилия, разнолистность, представляет собой явление, когда на одном растении развиваются как типично подводные листья, так и типично воздушные с рядом переходов (жеруха, стрелолист, поручейник). Погруженные листья могут сменяться плавающими, совершенно отличными от первых (виды сем. нимфейных, плавающий рдест).
6. Обилие воды, облегчая некоторые отправления растения, в тоже время грозит ему некоторой опасностью, именно подвергает растение риску выщелачивания из него необходимых ему веществ. Выделение слизи особыми железками препятствует выщелачиванию из растений питательных веществ , а также является защитой при временном пересыхании водоемов. Эта слизь в некоторых случаях, как например, у рогоза, может служить для уменьшения внутреннего трения органов растения друг о друга. Механическая ткань у водных растений, наоборот, редуцирована или не развивается вовсе, так как вода лучше поддерживает тело, чем воздух. Так как главное напряжение водным растениям приходится испытывать в виде натяжения, а не изгиба, то механическая ткань, где она имеется, соответственным образом сосредоточенна в виде центральны тяжей. Возможно, эта слизь имеет и бактерицидное защитное действие, подобно фитонцидам наземных растений.
7. Подавляющее большинство высших водных растений - многолетники. При перезимовывании часть видов целиком опускается на дно водоема, большинство зимует в виде корневищ, клубней или зимующих почек (турионов). Турионы морфологически представляют собой видоизмененные побеги, запасающие к осени питательные вещества, прежде всего крахмал, и погружающиеся на дно водоема. Весной, зимующие почки прорастают и всплывают на поверхность.
Таковы в общих чертах основные приспособления высших растений к обитанию в водной среде. Даже краткий перечень этих особенностей показывает, насколько велика жизненность видов, способных переносить значительные неблагоприятные изменения среды, приспосабливаясь к новым условиям. Наличие устойчивого механизма гомеостаза позволяет высшим водным растениям захватывать значительные территории и иметь широкое географическое распространение. Такие виды создают популяции, приспособленные к крайним условиям ареала, к значительным колебания света, температуры и др.
1.2 Приспособление растения к водной среде.
1. Для водных растений характерна гетерофиллия - развитие на одном и том же растении листьев типично водных (погруженных), плавающих или возвышающихся над поверхностью воды. Разнолистность характеризует большую пластичность листовой пластинки (и естественно самого растения) к изменяющимся условиям обитания. Так, у некоторых растений при прорастании под водой вначале образуются листья, приспособленные к водной среде (линейные у стрелолистов), затем появляются плавающие на водной глади и потом - воздушные. Эти два или три типа листьев резко отличаются друг от друга как морфологически, так и анатомически.
2. Центральное расположение механических элементов. У большинства сухопутных растений механические ткани и клетки сосредоточены главным образом ближе к периферии, а у водных растений ¾ в центре стебля, черешке, цветоножке, и в листьях по центральной оси листовых долей или среди мякоти в широких листьях. Благодаря такому строению тонкие и длинные цветоножки и черешки плавающих листьев приобретают прочность при волнениях или течениях.
3. Недостаток в воде воздуха приводит к сильному развитию воздухоносных полостей, поэтому у всех групп водных растений очень хорошо развита система межклетников, заполненных обычно воздухом. Эта система межклетников присутствует во всех органах водных растений. Эта воздухоносная ткань называется аэренхимой, служит источником кислорода и углекислого газа, необходимых для жизни растений. Она же уменьшает вес гидрофитов, помогая им удерживать на воде листья, цветки, а также вертикально располагаться в воде стеблям.
4. Поглощение воды и питательных веществ у водных растений происходит почти всей поверхностью листьев и стеблей. Для многих гидрофитов характерно слабое развитие корневой системы или полное её исчезновение. У многих видов корни служат в основном для закрепления растений на дне водоема, а корневища являются органом запаса пластических веществ и для вегетативного размножения (кувшинки, кубышки, рдесты, нимфейник, бразения, аир).
У водных растений хорошо развито вегетативное размножение. Отделенная или оторванная часть сальвинии способна существовать самостоятельно и разрастаться в новое растение. У других растений этой же группы образуются специальные вегетативные почки, которые предназначены для размножения и для перезимовки (водокрас, бразения, пузырчатка). Семенное размножение гидрофитов в сравнении с вегетативным менее надежно. У большинства водных растений после опыления цветок закрывается и опускается под воду, где и происходит созревание плода. Погружение стеблей и зимних почек возобновления осенью на дно происходит, видимо, благодаря накоплению в клетках крахмала, вследствие этого удельный вес увеличивается, и стебель или почка тонут. Весной крахмал превращается в растворимые углеводы и почка всплывает.
Немногочисленные водные однолетники размножаются семенами, которые часто снабжены специальными приспособлениями к удержанию их на поверхности воды, что способствует распространению семян на значительные расстояния.
У некоторых гигрофитов и гидрофитов имеются специальные клетки, выделяющие слизь, которая на отдельных частях погруженных растений может иметь разное значение в их жизни: препятствует быстрому высыханию, предохраняет подводные части от выщелачивания, уменьшает внутреннее трение органов растения. У бразении слизь способствует плавучести побегов, а зимой защищает вмерзающие в лёд верхушки побегов от низкой температуры и от возможной потери воды в сухой толще льда.
При изучении растительности водоемов исследователю приходится обращать внимание не только на растительность самого водоема, но часто и на растительность низменной части его берегов, особенно водохранилищ с колебаниями уровня. Таким образом, он имеет дело с растениями, разными по экологии, по их отношению к воде:
гидрофитами
- настоящими водными растениями, полностью или большей своей частью погруженными в воду;
гигрофитами
- растениями избыточного увлажнения;
мезофитами
- достаточного (среднего) увлажнения.
Водные растения являются вторично водными организмами - приспособившимися к жизни в водной среде наземными растениями. Виды их относятся к самым разнообразным и отдаленным друг от друга семейст. Большинство водных растений цветет и плодоносит над водой. Тех, у которых весь цикл развития совершается под водой, сравнительно немного. Кроме генеративного способа размножения, часто подавленного, у водных растений широко развито вегетативное размножение при помощи корневищ, частей стеблей, почек и т. д. Некоторые виды размножаются только вегетативным путем.
Глава 2.
2.Изучение флоры и фауны пруда.
2.1. ЭКОЛОГО-БИОЛОГИЧЕСКИЕ ГРУППЫ (БИОМОРФЫ)
ВОДНЫХ РАСТЕНИ
Вода является для рассматриваемой группы водных растений главной средой обитания. Важнейшие биологические особенности этой группы видов заключаются в приспособлении к жизни в воде или к условиям избыточного увлажнения. Провести четкую границу между настоящими водными растениями и группой прибрежно-водных растений часто очень трудно и поэтому понятие водное растение разными исследователями трактуется весьма различно (Белявская, 1982). Однако, несмотря на несколько отличные термины, общие признаки, характеризующие выделяемые группы растений, совпадают, так как многие авторы при разработке своих классификаций как основу используют классификацию А.П. Ренникова (1950), согласно которой различают следующие 3 группы высших водных растений:
I. Гидрофиты погруженные:
а) полностью погруженные в воду
(истинно водные) растения, весь цикл развития они проходят в воде: полностью погруженные неукореняющиеся
,
взвешенные (плавающие) в толще воды - виды рода Ceratophyllum
demersum (Роголистник погруженный)
и другие; и полностью погруженные укореняющиеся
-
виды родов Myriphyllum specatum(Уруть колосистая),
и другие;
б) погруженные в воду с воздушными генеративными органами
(почти погруженные):
1) погруженные неукореняющиеся
, взвешенные (плавающие) в толще воды - виды мы не нашли
2) погружен
ные укореняющиеся
, с различной мощности корневой системой (у некоторых иногда не развивающейся) - виды родов Potamogeton
L(рдест плавающий),Rorippa amphibia(Жерлянка земноводная)и другие.
II. Гидрофиты плавающие:
свободно плавающие неукореняющиеся
- Lemna
minor
L(ряска маленькая).,
lemna
trisulca
(ряска трехдольная),
Spirodela
polyrhiza
(L
.)
Schleid
(многокоренник)
и другие;
Погруженные и плавающие неукореняющиеся растения прикрепляются к субстрату в тех случаях , когда нижние части их стеблей или водных корней находятся в рыхлой иловой толще дна водоема.
III. Воздушно-водные гелофиты - водно-болотные растения
Надводные растения с поднимающимися (возвышающимися, выставляющимися) над поверхностью воды стеблями и листьями, укореняющиеся - (typha latifolia) рогоз широколистныйи другие.
Они успешно существуют и проходят полный цикл развития, как в воде, так и на влажных берегах водоемов.
2.1 Изучение флоры.
Изучение флоры проводили с 23 июня по 15 июль 2008 года.. Маршруты проходили по берегу и по водной поверхности на лодке. Собирали растения для гербария и с помощью водного сачка ловили животных обитателей пруда. Затем с помощью определителя классифицировали собранный материал. В процессе работы были сделаны фотографии водных растений.
В аннотированном списке даны русские и латинские названия семейств и видов, экологическое произрастание, экологическая группа.
Были проведены систематический и экологический анализы. Особое внимание было уделено анализу популяций водных растений. .
Список водных растений.
1
. Сем. Рдестовые (Potamogetonaceae
Dumort), род Рдестовые
вид Рдест плавающий (Potamogeton
natans
L).
2.
. Сем. Осоковые (Cyperaceae
Juss)
1) род Осока, вид Осока водяная.(Carex
aquatilis
)
2)род Камыш вид Камыш озёрный(scirpus
lacustris
)
3
Сем. Рясковые ( Lemnaceae
S. F. Gray)
род Многокоренник, вид Многокоренник обыкновенный (Spirodela
polyrhiza
(L
.)
Schleid
)
.
2)род Ряска, вид Ряска трехдольная (lemna
trisulca
),
3) вид Ряска маленькая(lemna minor
),
4.
Сем. Роголистниковые,род Роголистник, вид Роголистник погруженный
(ceratophyllum
demersum
)
5.
Сем. Сланоягодниковые, род Уруть, вид Уруть колосистая (Myriophyllum
specatum
)
6.
Сем. Крестоцветные, род Жерушник, Жерушник земноводный(rorippa
amphibia
)
Прибрежные растения
7
Сем. Гераниевые, род Герань, вид Герань луговая(Geranium prtense)
8.
Сем. Лютиковые,род Лютк, вид Лютик едкий(Ranunculus repens)
9
.Сем. Маренные, род Подмаренник вид Подмаренник мягкий,(gallium mollugo)
10.
Сем. Подорожниковые- (Plantaginaceae
Juss).
род Подорожник, вид Подорожник песчаный-(Plantago
arenaria
Waldst. et Kit).
11.
Сем. Колокольчиковые- (Campanulaceae
Juss).
Род Колокольчик, вид Колокольчик персиколистный;
12.
Сем.Сложноцветные,род Тысячелистник, вид Тысячелистник обыкновенный
(Achillea millefolium)
13
.Сем. Сложноцветные:
1)род Цикорий, вид Цикорий обыкновенный,(Cichorium intybus)
2) род Пижма, вид Пижма обыкновенная или дикая рябина(Tanacetum vulgare)
3)род Одуванчик, вид Одуванчик лекарственный(Taraxacum officinale)
4)род Мать – и – мачеха, вид Мать – и – мачеха обыкновенная
14
Сем. Злаки:
1)род Мятлик, вид Мятлик луговой(Poa pratensis)
2) род Ежа, вид Ежа сборная(Dactylis glomerata)
15.
Сем. Зверобойные, род Зверобой, Зверобой продырявленный (Hypericum perforatum)
16. Сем. Хвощовые, род хвощ, вид Хвощ приречный (Equisetum fluviatili L.)
17. Сем. Бобовые, род Чина, вид Чина болотная (Lathyrus palustris L.)
2.2. ЭКОЛОГО-БИОЛОГИЧЕСКИЙ АНАЛИЗ
В зависимости от отношения растений к фактору увлажнения и по степени приспособленности их к жизни в водной среде растения исследованной территории можно объединить в три группы (табл. 3).
Экологическая группа ¾ гидрофиты погруженные представлена одним полностью погруженными укореняющимися видом роголистник погруженный (Spirodela
polyrhiza
(L
.)
Schleid
)
.
Роголистник может расти на большой глубине, до 9 метров. У растения длинный стебель, ветвящийся в верхней части. У растения длинный стебель, ветвящийся в верхней части. Листья на стебле расположены в мутовках. Листья темно-зеленые, жесткие, хрящевинные. Они сильно рассечены на узкие, как ниточки, части- сегменты, часто мелкопильчатые по краю. Цветки роголистника мелкие, едва заметные, без лепестков. Тычиночные и пестичные цветки расположены на одном растении. Корней у роголистника – нет. Их роль выполняет особые бледные, почти бесцветные ветви в . Они проникают в ил и, как якорь, удерживают растение. Питательные (минеральные) вещества поглощает вся поверхность роголистника: и стебель, и листья, а не корни, как у большинства растений. ¾ и тремя видами с воздушными генеративными органами ¾ Ряска трехдольная (lemna
trisulca
)ряска маленькая (lemna minor
), и многокоренник обыкновенный
- (Spirodela
polyrhiza
(L
.)
Schleid
)
.занимает достаточно мелководные участки (0,3-0,9 м). Кроме того, в эту экологическую группу входят пять видов укореняющихся растений с плавающими листьями: уруть колосистая.
У этого растения длинный, до полутора – двух метров, стебель. На нем мутовками по 4-6 штук сидят сильно рассеянные перистые листья. Перистыми их называют за некоторое сходство с птичьим пером: от центральной «ости» отходят вбок многочисленные тонкие сегменты листа. Растет уруть в основном в стоячей воде. Ее также можно встретить в озерах, тихих речных заводях, в старицах рек. Рдест плавающий.
Рдесты – многолетние водные растения. В грунте у них развивается длинное корневище. Оно перезимовывает, а весной из зимующих почек отрастает удлиненные побеги. Отдельные побеги или их часто могут отрываться и свободно плавать в воде, продолжая свое развитие. Рдестами питаются водные моллюски, насекомые, рыбы. В зарослях рдестов многие рыбы мечут икру. Отмершие побеги падают на дно. Разлогаясь, они превращаются в плодородный ил. Рдест плавающий – обычно растение озёр, прудов, речек. Предпочитает медленно текущую воду. Колосовидное соцветие зеленовато, возвышается над водой, цветет в июне – августе.
В эту экологическую группу входят наиболее декоративные растения, листья которых покрывают частично водную поверхность пруда, а цветки являются дополнительным украшением на пруде.
Наибольшее количество растений входит в группу гелофитов, то есть являются водно-болотными растениями ¾ вида. Эти растения произрастают вдоль берега пруда. Доминирующими по занимаемой площади здесь являются виды рода Carex
(Carex
aquatilis
),
образуя крупные кочки, обрамляющие пруд.
Таблица 3
Экологические группы растений биоценоза « Рыбный пруд»
Экологическая группа ¾ гидрофиты, живущие в воде:
Обыкновенный прудовик
- один из самых крупных прудовиков нашей страны. Раковина его спирально закручена, имеет острую вершину из 4-5 оборотов, которая достигает в высоту 45- 65 мм, а в ширину 27 мм. Он живет в небольших озерах , затонах рек в небольших прудах, заросших растениями.
Пиявка ложноконская большая
. Ложноконская пиявка имеет длину тела до 15см, в ширину достигает 15 мм. Окраска спины взрослой пиявки черная с коричневым оттенком и темными пятнами; спина молодых особей более светлая с узором, брюхо зеленовато-серое. Имеет 5 пар глаз, а в ротовой полости - челюсти с острыми краями. Ложноконская пиявка обитает в водоёмах со стоячей или медленно текущей водой: в равнинных реках, озерах, прудах, иногда даже в лужах. Она и прибрежных в зонах озер. Передвигается в водоеме разными способами: может ползать и плавать.
Обыкновенный карась.
Обыкновенный карась - одна из самых известных рыб, длина тепа до 45 см, а масса достигает 3 кг. Тело высокое, сжатое с боков. Цвет спины темно – золотистый, бока темно – зеленые, брюхо светлое. Парные плавники красноватые. Карась живет в заболоченных, заросших водной растительностью водоемах, в местах с замедленным течением и илистым грунтом. Эта неприхотливая рыба может существовать при минимальном содержании кислорода в воде; хорошо переносит и холодную зиму.
Паук- серебрянка
- водяной паук. Размеры самца 14- 17 мм, самки 11-12ммОкраска головогруди ржаво – красноватая с темным рисунком; брюшко – бурое, покрыто светло – серыми волосками. Паук – серебрянка широко распространено по всей территории нашей страны. Он обитает в стоячих или медленно текущих водоемах, богат растительностью. Паук – серебрянка питается животной пищей: рачками, личинками насекомых, мальками рыб.
Водяной скорпион.
Водяной скорпион - один из водных клопов Длина его тела 18-22 мм, дыхательная трубочка около 11мм. Окраска верха тела серо - бурая, бока рыжие. Передние ноги похожи на клешни. Водяной скорпион живет на дне прудов, а также рек с медленным течением, заросших воной растительностью. Плавать этот клоп не умеет. Он либо сидит на растениях, либо ползает между ними. Его трудно заметить из-за невзрачной бурой окраски. Водяной скорпион – хищник, он подкармливает мелких животных: личинок насекомых, червей, мальков рыб.
Прудовая
лягушка
(Rana lessonae
Camerano, )
Сверху лягушка ярко-зеленого, серо-зеленого цвета, с темными пятнами . Вдоль спины проходит светлая продольная полоска, снизу чисто-белого или желтоватого цвета. Самцы с белыми резонаторами в углах рта. Обитает в водоемах смешанных и широколиственных лесов, после размножения можно встретить далеко от воды. Активна днем, ночь проводит на дне водоема. Кормится жуками, стрекозами, муравьями. Зимняя спячка длится 100 дней. Икру мечет в мае, не большими порциями. Самка откладывает до 3000 яиц. Развитие личинок 70-135 дней. Зрелой лягушка считается на третий год. Включена в Красную книгу Удмуртии (Красная книга УР, 2001).
Озерная лягушка (Rana
ridibunda
Pall.,)
Спинка буровато-зеленая с темными пятнами. Вдоль спины проходит светлая продольная полоса. Снизу грязноватого или желтоватого цвета с темными пятнами. Обитает в водоемах, быстрых речках. Всю жизнь проводит в воде или недалеко от нее. Активна круглые сутки. Кормится насекомыми, изредка схватывает птенцов мелких птиц, полевками, головастиками, мальками рыб. На зимовку уходит на дно водоемов в сентябре, выходит в июне. Количество яиц до 3989. Икру откладывают в виде двух больших комков или группами по 8-10 икринок. Головастики вылупляются через неделю, длина головастика 50-90 мм. Зрелой лягушка считается на третий год.
Жерлянка
.Жерлянка похожа на небольшую жабу, сверху сухая или черная, снизу оранжевая, в черных пятнах. Минорные и однозвучно – мелодичные голоса жерлянок навевают грусть. Самец – жерлянка лежит, распластанный, на поверхности пруда. Он кричит «унк», и тело его вибрирует. По воде мелкой дрожью разбегаются круги. При теплой погоде таких «унк» раздается 18 – 20 в минуту. Кожа жерлянок ядовита. Не так смертельно, как, скажем, древолазов, но сильнее, чем у всех наших жаб. Предупреждает о своей несъедобности жерлянка так: переворачивается черно – красным брюхом вверх. Когда напугана, словно пеной, покрывается ядовитыми выделениями.
На поверхности воды
Водомерка прудовая
- небольшой клоп длиной 8-10 мм. Грудь сверху черна, на боках светлая продольная полоска. Бедра передних ног светлые с черными продольными полосками . Она обитает на поверхности водоемов (прудов рек). Кормится прудовая водомерка мелкими насекомыми, падающими на поверхность воды. Для захвата пищи используется короткие передние ноги
Животные, обитающие около пруда
Синяя стрекоза
- это насекомые с двумя парами сильных перепончатых крыльев, с помощью которых они могут летать с большой скоростью. Часто крылья окрашены в яркие цвета, и это придает насекомому необычайную привлекательность. Стрекозы являются хищниками. Они ловят добычу на лету, хватая ее своими мощными челюстями. У стрекоз очень большие глаза, и они способны заметить жертву с большого расстояния.
Большое коромысло
- одна из крупных стрекоз; длина ее брюшка достигает 49- 60 мм, крыльев- 45- 49мм. Размах крыльев до 10 см. Крылья желтоватые, прозрачные, с бурыми жилками; бока рыжевато – бурые с желтыми полосами. Она живет недалеко от водоёма, но нередко ее можно встретить на опушке леса или на лугу, расположенный не значительно расстояние от водоема. Большое коромысло активна с середины лета до поздней осени , предпочитает солнечные жаркие дни.
ОНДАТРА, или
МУСКУСНАЯ КРЫСА
Длина тело ондатры 25-35см, длина хвоста 27-30см; масса до 1,5 кг. Ушная раковина едва выступает из меха. Хвост с боков покрыт редкими волосами и мелкими чешуйками. Пальцы задних лап соединены небольшой плавательной перепонкой. Окраска верха от темно-рыжей до темно - бурой, низа - более светлая. Ондатра – североамериканский вид, который в настоящее время акклиматизирован почти, но всей территории нашей страны.
2.4. ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ ИССЛЕДОВАНИЯ
В результате проведенной работы нами отмечены на пруде 24 вида сосудистых растений из 23 родов и 17 семейств. Ведущее положение занимают семейства Сложноцветные(5видов) , семейства Злаки (2вида) и семейства Рясковые (3вида). остальные семейства включают по одному по - одному.
В зависимости от отношения растений к фактору увлажнения и по степени приспособленности их к жизни в водной среде растения исследованной территории были объединены нами в три группы.
Наибольшее количество растений входит в группу гелофитов, то есть являются водно-болотными растениями ¾ 3 вида. Эти растения произрастают вдоль берега озера, на заболоченных участках, примыкающих к озеру. Доминирующими по занимаемой площади здесь являются виды рода рясок: ряска трехдольная (lemna trisulca),ряска маленькая (lemna minor), и многокоренник обыкновенный - (Spirodela polyrhiza (L.) Schleid)..
Истинно водные растения входят в экологические группы ¾ гидрофиты погруженные и гидрофиты плавающие.
Роголистник погруженный (ceratophyllum demersum) относятся к полностью погруженным укореняющимися гидрофитам, весь жизненный цикл которых проходит в воде.
А так же нами было изучено: 3 вида амфибий, 1 вид млекопитающего,1 вид рыбы 5 видов водных обитателей(прудовик, паук – серебрянка) и 2вида стрекоз.
Выводы
Водно - прибрежные растения рыбного пруда представлены 19 видами цветковых растений из 18 родов,12 семейств, а так же 6 видов из 5 родов и семейств водных растений.
Нами были исследованы самые распространенные виды насекомых и животных, как например: ондатра, большое коромысло и. т.д.
ЛИТЕРАТУРА
1. Катанская В.М. Высшая водная растительность континентальных водоемов СССР. Методы изучения. Л.: Наука, 1981. 187с.
2. Кокин К.А. Экология высших водных растений. М.: Изд-во Моск. Ун-та, 1982. 160с., ил.
3. Шенников А.П. Экология растений. М.: Совет. Наука, 1950. 375с.
4. Крюкова М.В. Водно-прибрежная флора Среднеамурской низменности: Автореф. дис... канд.биол. наук. Владивосток, 2000. 24 с.
5. Михайловская И.С. Строение растений в связи с условиями жизни . М.: Моск. гос. заочн. пед. ин-т, 1964. 92 с.
6. Пашкевич В.Ю., Б.С. Юдин. Водные растения и жизнь животных. Новосибирск: Наука, 1978. 128 с.
7. Рычин Ю.В. Флора гигрофитов. М.: Советская наука, 1948. 448 с.
Понятие о гидрофитах.
Достаточно давно водные растения стали называть гидрофитами или макрофитами.
Последний термин часто служит синонимом термина «высшие водные растения». Однако многие специалисты считают, что под термином «макрофиты» целесообразно понимать макроскопические, т.е. доступные наблюдению невооруженным глазом, растительные организмы вне зависимости от их систематического положения (Распопов, 1985). В этой связи к категории макрофитов наряду с высшими растениями часто относят харовые водоросли, скопления многих нитчатых водорослей (например, виды родов С1аdophora, Rhizoclonium, Spirogyra
и др.).
Современную высшую водную растительность, обитающую в пресных, солоноватых и в значительно меньшей мере морских водоемах, принято называть вторично-водной, т.е. это потомки более древних наземных растений, приспособившихся к жизни в воде. Общую схему эволюционного пути водных растений можно представить следующим образом: море -> пресные воды -> суша -> пресные воды -> море (Кокин, 1982).
Экологические особенности водной среды.
Распространение высших водных растений зависит от многих факторов внешней среды: света, содержания растворимых газов, температуры и биогенных веществ и др.
Световой режим
в воде отмечается заметным ослаблением освещенности и изменением спектрального состава света с глубиной. Например, в самых чистых водах Саргассова моря прозрачность достигает глубины 66,5 м (величину прозрачности определяют путем погружения в воду белого диска диаметром около 30 см до предельной глубины его видимости). В то же время в мутных проточных водах уже на глубине 50 см количество света может уменьшаться до 7%, т.е. до такой же величины, как и под густым пологом елового леса.
Газовый режим
водоемов существенно отличается от назем-но-воздушной среды, и прежде всего меньшим количеством воздуха и иным его составом. Так, содержание углекислого газа в воде на единицу объема при одинаковой температуре почти в 700 раз больше, а кислорода в 30-35 раз меньше, чем в воздухе. Причем если в воздухе концентрация кислорода достаточно постоянна (21%), то в воде его содержание значительно меняется. Это может быть дефицит кислорода (ночью или зимой подо льдом) или повышенная концентрация в дневное время в случае массового размножения фитопланктона.
Температурный режим
водоемов характеризуется прежде всего отсутствием резких перепадов температуры в течение года и суток. Это связано с высокой удельной теплоемкостью и теплопроводностью воды, которые в 500 и 30 раз соответственно выше, чем у воздуха. Именно поэтому вода медленно нагревается и медленно остывает. В результате амплитуда суточных и сезонных колебаний температуры в водоемах относительно низкая.
Солевой режим
гидросферы определяется содержанием в воде разнообразных минеральных веществ - карбонатов, сульфатов, хлоридов. В зависимости от количества растворимых веществ выделяют пресные
(до 0,5 г/л), солоноватые
(0,5 -16 г/л), морские
(16-47 г/л) и пересоленные
(47 - 350 г/л) воды. Для гидробионтов наибольшее значение имеют хлориды натрия, магния и серы, а также сульфат магния, растворенные в воде.
Основными элементами минерального питания макрофи-тов являются азот, фосфор, кремний, железо, марганец, которые встречаются во всех пресных водоемах в виде нитратов, фосфатов, кремнезема и др.
Экологические особенности водных растений.
Экологические особенности водной среды накладывают определенный отпечаток на морфолого-анатомические и физиолого-биохимиче-ские признаки гидрофитов. Важнейшими чертами структурно-функциональной организации макрофитов являются следующие.
1. Большинство водных растений - корневищные многолетники, способные расти не только в воде, но и на суше. Это выражается в явлении диморфизма, когда при высыхании водоема растения не погибают, а продолжают жить, изменившись морфологически в наземную форму. Таковы некоторые виды шелковников (Ваtrachium),
стрелолистов (Sagittaria),
калужниц (Caltha),
горец земноводный (Реrsicaria amphibia)
и др. Однолетних видов среди водных растений мало. Многие из них цветут и плодоносят под водой.
2. В связи с малым количеством света в воде и иным его спектральным составом для макрофитов характерно большое развитие поверхности тела по отношению к его массе. Это достигается за счет ветвления тонких длинных стеблей и рассечения листовых пластинок на узкие тонкие полоски (роголистники (Ceratophyllum),
урути (Муriophyllum),
шелковники и др.). Увеличение поверхности листьев позволяет водным растениям полнее использовать свет и СО 2 в процессе фотосинтеза. Доказательством тому, что это действительно является важным приспособительным признаком, служит явление гетерофиллии
(разнолистности), т.е. наличие на одном побеге листьев разной формы - подводные листья сильнорассеченные, а надводные менее рассеченные или цельные (рис. 6.1). Гетерофиллия хорошо выражена у кувшинок (Nymphaea)
и кубышек (Nuphar),
стрелолиста, поручейника широколистного (Sium latifolium)
и у некоторых других видов.
3. У водных растений наблюдается частичное или полное отсутствие дифференцировки мезофилла на палисадный и губчатый, что связано с резким уменьшением доступа света к ним.
4. В связи с большой плотностью воды у макрофитов механическая ткань развита слабо или совсем отсутствует. Механические элементы у ряда видов, обеспечивающие гибкость стеблей и листьев при сильном течении или волнении воды, расположены в центральной части стебля и по центральной оси листа, обычно вокруг проводящих пучков.
5. У многих водных растений в клетках эпидермиса имеются хлоропласты, что способствует более эффективному использованию света.
6. Проводящие элементы ксилемы развиты слабо. Поглощение воды и растворенных в ней веществ происходит всей поверхностью погруженных в воду частей тела растения, что сопровождается некоторой редукцией проводящей ткани.
7. У макрофитов хорошо развита воздухоносная ткань -аэренхима, образованная системой межклетников, пронизывающая все тело растения, что обеспечивает более эффективный газообмен.
8. Листья погруженных водных растений часто лишены устьиц; газообмен совершается всей поверхностью тела. В то же время у водных растений с плавающими листьями устьица расположены на верхней стороне листа и их количество в сравнении с наземными растениями увеличено.
9. Для макрофитов характерна редукция корневой системы или изменение ее функции. Так, корни многокоренника (Spirodela polyrhiza)
и рясок (Lemna) -
это орган равновесия, а корневая система водяного ореха (Тrapa natans)
служит для прикрепления их к грунту и выполняет функцию фотосинтеза.
10. У водных растений вегетативное размножение преобладает над семенным. Самый распространенный способ зимовки многолетних растений - в виде побегов с сильно укороченными междоузлиями или в виде особых зимующих почек, образующихся на материнском растении в конце вегетационного периода.
11. Для водных растений характерен усиленный рост по сравнению с наземными, поскольку в течение сравнительно короткого вегетационного периода растение должно пройти весь цикл развития.
12. Плоды и семена многих макрофитов долго сохраняют способность к прорастанию.
Таковы в общих чертах основные приспособления высших растений к обитанию в водной среде. Наличие генетически обусловленного устойчивого механизма гомеостаза позволяет гидрофитам осваивать значительные территории и иметь широкое географическое распространение.
Классификация водных растений.
В качестве естественной биологической группы водные растения стали рассматриваться лишь в конце XIX в. Согласно одной из первых сводок, посвященных описанию жизни пресных вод (Ламперт, 1900), водные растения подразделены на следующие группы:
1) растения с листьями, погруженными в воду;
2) растения с листьями, плавающими на поверхности воды;
3) растения со стеблями и листьями, частично погруженными в воду и частично «выставляющимися из воды в воздух».
Все последующие многочисленные варианты классификации водных растений и водной растительности, разработанные различными авторами, в том или ином виде включают в себя эти основные группы.
Наиболее часто используются классификации Г.И. Поплав-ской (1948) и А.П. Шенникова (1950). Г.И. Поплавская выделяет две группы водных растений: гидрофиты -
растения, меньшей своей частью погруженные в воду, и гидатофиты -
растения полностью или большей частью погруженные в воду. Гидатофиты в свою очередь она подразделяет на три группы: гидатофиты настоящие, аэрогидатофиты погруженные и аэрогида-тофиты плавающие.
А.П. Шенников, напротив, к группе гидрофитов относит погруженные в воду растения и растения с плавающими листьями или листоподобными стеблями. Воздушно-водные растения отнесены им к гелофитам.
К водным растениям (гидрофитам) близки гигрофиты -
исконно сухопутные растения, приуроченные к избыточно увлажненным местообитаниям. Поскольку многие виды гигрофитов по своим морфолого-анатомическим особенностям схожи с гидрофитами, в последние годы наблюдается тенденция включения в списки флоры водоемов большого числа прибрежных (околоводных) растений.
По мнению Г.С. Гигевич, Б.П. Власова, Г.В. Вынаева (2001), «с таким подходом нельзя согласиться, поскольку в водной среде по самым разным обстоятельствам, например, при создании искусственного водоема, при половодье, обвале части берега в воду и т.п., типично сухопутные виды растений могут оказаться в воде. С другой стороны, не всегда можно с уверенностью сказать, водные это растения или сухопутные. Многие гидроботаники, например, рассматривают в качестве водных растений ряд представителей рода осока (Сагех), растущих по берегам водоемов, хотя в большинстве своем эти осоки скорее обитатели сырых, чем покрытых водой грунтов».
Для того чтобы прийти к какому-то единому мнению по этому вопросу, пожалуй, целесообразно присоединиться к мнению известного российского гидроботаника И.М. Распопова (1978, 1985) о том, что «.высшими водными растениями, по-видимому, следует считать травянистые растения, анатомически и морфологически приспособленные к жизни в водной среде в погруженном, плавающем на поверхности или полупогруженном состоянии, и за ними целесообразно закрепить название "гидрофиты"».
В настоящее время не существует единой общепринятой лассификации водных растений, поскольку гидрофиты не представляют собой единого целого. Из многочисленных классификаций гидрофитов наибольший интерес представляет экобиоморфологическая система В.М. Катанской (1981), согласно которой все водные растения по своим морфологическим (формы роста) и эколого-биологическим особенностям объединяются в следующие экологические группы.
Hygrophila corymbose,
с подводными и надводными побегами
в одной из рек на Сулавеси
Цветущая кувшинка (Nymphaea lotus)
в одном из временных водоемов
в Сенегале
Ceratopteris pteridoides
формирует утолщенные губчатые
черешки листьев, повышающие
плавучесть растений,
р. Рио-Янаяку, Перу
Для многих тропических и субтропических болотных растений, которые произрастают во временных или затопляемых районах в водоемах со стоячей или проточной водой, характерна способность приспосабливаться к периодической смене дождливого и засушливого сезонов.
Растение в засушливый период растет сравнительно близко от водоема, либо вне воды, либо на несколько сантиметров в воде. При полном высыхании водоема это может привести к полному отмиранию внешних частей растения. В период полой воды, наоборот, они ведут частичный или полностью подводный образ жизни. Таким образом, растения подобной среды обитания при смене сезонов в состоянии приспосабливаться к подводному или надводному образу жизни. У них перемежающийся жизненный цикл, и они должны за определенный период, от начала дождей и до вероятного отмирания в конце засушливого сезона, (отцвести и принести семена) чтобы гарантировать дальнейшее существование вида.
При наступлении сезона дождей у многих водных и болотных растений сначала формируются характерные ювенильные (юношеские) формы. Из сохранившихся в почве в период засухи вегетативных органов (споры, клубни, луковицы) они вначале образуют слабенькие листья, которые бывают узкими, мягкими и прозрачными. При прорастании семян после формирования одного или двух зародышевых листочков обычно появляются эти молодые листики. Только с возрастом и ростом растений формируются присущие каждому виду подводные и плавающие листья. Последние своим анатомическим строением, характеризующимся отсутствием устьиц, тонким слоем эпидермиса и обширными воздушными каналами, особенно хорошо адаптируются в водной среде. Благодаря этому водные листья способны усваивать кислород, углекислоту и питательные вещества непосредственно из воды. В сухом воздухе листья быстро увядают и засыхают из-за незначительности механической ткани и слабой защиты от испарения. Как только уровень воды снова начинает понемногу спадать, эти земноводные растения переходят в свое надводное состояние, то есть у них уже не формируются подводные листья, а укрепившиеся наружные листья выбрасываются над водной поверхностью и вырастают в полный размер. Эти надводные листья у многих видов имеют совсем иной облик, нежели подводные (гетерофиллия). Если подводные листья часто нежные, тонкие, прозрачные и лентообразные или рассеченные, что увеличивает их поверхность, то характерными признаками наружных листьев являются цельные края, жесткая, кожистая и часто опушенная поверхность. Образование листьев разной формы в различных средах особенно хорошо наблюдается у Нуdrоphila difformis
, так же, как и у разнообразных видов Limnophila
и Myriophyllum
. Вновь снижающийся уровень воды снова прекращает цветение и образование семян. Как только биотоп начинает пересыхать, растения приобретают все более грубый габитус, вплоть до того, что от снижения влажности почвы у него полностью отсыхают листья. Если у некоторых растений в пересохшей почве все же сохраняются органы, способствующие выживанию, то другие виды полностью отмирают, и лишь их семена пребывают в состоянии покоя, пока не наступит новый сезон дождей и они благодаря благоприятным условиям не начнут прорастать. У многих настоящих водных растений при достижении ими водной поверхности формируются плавающие листья, которые плоско ложатся на поверхность воды и анатомически приспосабливаются к воде. У них особенно развиты межклетники, способствующие фотосинтезу. Одновременно плавающие листья некоторых видов, например кувшинки, обладают множеством гидропотов, (особые клетки эпидермиса) на тыльной стороне листа, с помощью которых они могут усваивать воду и питательные минеральные вещества. Подобные гидропоты имеются и на листьях множества водных растений.
Некоторые такие растения, как кувшинка (Nymphaea
) и кубышка (Nuphar
) способны с помощью своих корней и корневищ давать побеги в плохо вентилируемой, бедной кислородом почве. В качестве морфологической адаптации к подобной среде они сформировали систему полостей, с помощью которой части растения, находящиеся в земле, могут снабжаться кислородом от листьев, плавающих на воде. Эти полости настолько велики, что, например, через черешок листа кувшинки можно продувать воздух.
В особенно плотных почвах у растений тропических водоемов иногда можно встретить корни, растущие поверх земли, корни, образующие побеги, и в качестве сверхприспособляемости к такой среде обитания - дыхательные корни. Формирование таких дыхательных корней хорошо известно у некоторых видов людвигии, которые образуются главным образом в бедной кислородом среде и отличаются от обыкновенных корней наличием белой, губчатой ткани с межклеточными пространствами (аэренхима), предназначенной для вентиляции и запаса воздуха. Эти дыхательные корни растут вертикально вверх, вероятно поглощают кислород из атмосферы и через вентиляционную ткань подводят его к подводным побегам.
Плавающие растения имеют морфологические и анатомические приспособления. Так, например, у видов сальвинии оба плавающих листка снабжены множеством межклеточных пространств, в то время как опускающиеся в воду листья делятся на множество нитевидных долей, густо покрытых волосками, выполняющих функцию отсутствующих корней. Особенно обращают на себя внимание утолщенные губчатые черешки листьев у Ceratopteris pteridoides
(водяная капуста), Eichhornia crassipes
и Trapa natans
, которые богато снабжены вентилирующей тканью и тем самым облегчают растению плавание по поверхности воды. У Limnobium
тыльная сторона листа покрыта толстым губчатым слоем, что также повышает плавучесть растения.
У плавающих растений есть разнообразные способы защиты от смачивания для сохранения сухой от дождя и росы верхней транспирирующей части листа, находящейся над поверхностью воды. Хорошо известна несмачиваемость листов лотосов (Nelumbo nucifera
), усеянных бесчисленными бугорками. Но и у видов сальвинии плавающие листья надежно защищены от смачивания сосочками, расположенными рядами, которые в свою очередь покрыты волосками. Одновременно эти плавающие листья в форме лодочки способствуют быстрому удалению дождевых капель. У других видов плавающих растений (например, у Pistia stratiotes
) несмачиваемость достигается за счет множества волосков. Примечательна и выпуклая форма плавающих листьев некоторых водных растений (например, Phyllanthus fluitans
), которая также предназначена для быстрого удаления дождевой воды с поверхности.
Наличие лентообразных, гладких, волнистых или лишенных мякоти листьев, например, у некоторых видов Aponogeton
, Cryptocoryne
и Vallisneria
, является отличительными признаками некоторых водных растений. Подобную форму и структуру листьев можно объяснить как приспособительными, так и защитными функциями, поскольку листья вследствие своей особой формы или структуры поверхности оказывают наименьшее сопротивление сильному течению. Листья узкой или ленточной формы образуются в особенности у видов, живущих в проточной воде. Такая форма характерна и для реофитов. При этом речь идет о растениях, встречающихся в промежуточных между высокой и низкой зонах и которые на короткое время затапливаются. Листья у них жесткие, кожистые или грубые и имеют соотношение длины к ширине минимум 4:1.
У некоторых водных растений озер Малави и Танганьика можно наблюдать дополнительные адаптивные свойства. Оба африканских озера тектонического происхождения хорошо известны аквариумистам благодаря богатству форм своей ихтиофауны. В переходной зоне от осыпей к песчаному грунту, в зарослях тростника, а также в литоральной зоне встречаются растения, которые приспособились к особым условиям этих водоемов благодаря созданию специальных адаптивных форм. Так, у Vallisneria spiralis var. denseserrulata
удивительно короткие и очень жесткие листья, благодаря чему растения способны оказывать сопротивление волнообразным движениям. У Ceratophyllum demersum
и Myriophyllum spicatum
в этих озерах благодаря коротким побегам, а также очень жесткой структуре листа и стебля совершенно отличный от популяций других регионов обитания внешний вид, что можно объяснить приспособляемостью и защитой от сильных волн. Вдобавок у С. demersum
побеги так компактны, что не плавают свободно в воде, как это наблюдается у других популяций, а вследствие своей большой плотности опускаются на дно, где подвергаются меньшему волнению, чем на водной поверхности.
Оптимальной приспособляемости в естественной среде обитания достигает и гигантская лилия, водяная виктория амазонская (Victoria amazonica
). У растений огромные, до 2,5 метров в диаметре плавающие листья, на поверхности которых для газообмена и ассимиляции расположено необходимое количество устьиц. У этих плавающих листьев сильно развито жилкование, а также высокая, до 10 см, боковая кромка, отчего волнение встречает сильное сопротивление и лист не повреждается.
Другими приспособительными средствами к жизни в воде являются ловушки для животных у Aldrovanda vesiculosa
и ловчие пузырьки у видов Utricularia
, с помощью которых растения ловят мелких животных, переваривают и используют в качестве органического азота. Формирование зимних почек (турионов) следует рассматривать как приспособление многочисленных водных растений к неблагоприятным периодам вегетации. У многих водных растений формируются подобные почки, которые осенью опускаются на дно водоема, где и пережидают холодные времена. Следующей весной они вновь поднимаются на поверхность и дают новые побеги.
В этом контексте интересно и поведение рясковых (Lemnaceae
). Поздней осенью у них формируются измененные листецы с сильно развитой межклеточной структурой и большим содержанием крахмала, отчего повышается их удельный вес. Вследствие этого растения утрачивают свою плавучесть и падают на дно. Весной элементы вновь начинают расти и поднимаются на поверхность воды.
Помимо названных форм, приспособленных к жизни в воде, далее будут упомянуты многообразные механизмы опыления водных растений.
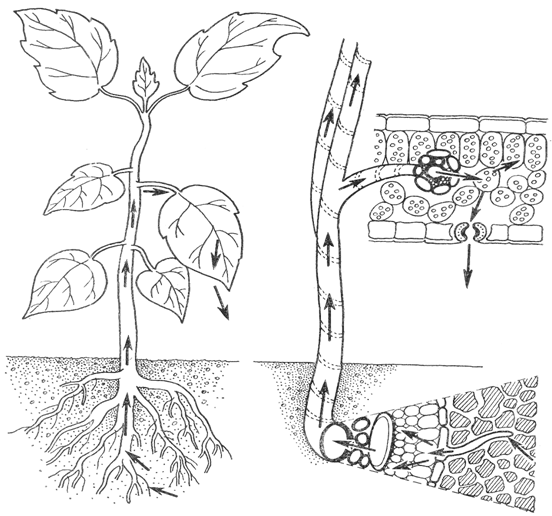



 Паук- серебрянка
- водяной паук. Размеры самца 14- 17 мм, самки 11-12ммОкраска головогруди ржаво – красноватая с темным рисунком; брюшко – бурое, покрыто светло – серыми волосками. Паук – серебрянка широко распространено по всей территории нашей страны. Он обитает в стоячих или медленно текущих водоемах, богат растительностью. Паук – серебрянка питается животной пищей: рачками, личинками насекомых, мальками рыб.
Паук- серебрянка
- водяной паук. Размеры самца 14- 17 мм, самки 11-12ммОкраска головогруди ржаво – красноватая с темным рисунком; брюшко – бурое, покрыто светло – серыми волосками. Паук – серебрянка широко распространено по всей территории нашей страны. Он обитает в стоячих или медленно текущих водоемах, богат растительностью. Паук – серебрянка питается животной пищей: рачками, личинками насекомых, мальками рыб.

 Прудовая
лягушка
(Rana lessonae
Camerano, )
Прудовая
лягушка
(Rana lessonae
Camerano, )
 ерлянка
.Жерлянка похожа на небольшую жабу, сверху сухая или черная, снизу оранжевая, в черных пятнах. Минорные и однозвучно – мелодичные голоса жерлянок навевают грусть. Самец – жерлянка лежит, распластанный, на поверхности пруда. Он кричит «унк», и тело его вибрирует. По воде мелкой дрожью разбегаются круги. При теплой погоде таких «унк» раздается 18 – 20 в минуту. Кожа жерлянок ядовита. Не так смертельно, как, скажем, древолазов, но сильнее, чем у всех наших жаб. Предупреждает о своей несъедобности жерлянка так: переворачивается черно – красным брюхом вверх. Когда напугана, словно пеной, покрывается ядовитыми выделениями.
ерлянка
.Жерлянка похожа на небольшую жабу, сверху сухая или черная, снизу оранжевая, в черных пятнах. Минорные и однозвучно – мелодичные голоса жерлянок навевают грусть. Самец – жерлянка лежит, распластанный, на поверхности пруда. Он кричит «унк», и тело его вибрирует. По воде мелкой дрожью разбегаются круги. При теплой погоде таких «унк» раздается 18 – 20 в минуту. Кожа жерлянок ядовита. Не так смертельно, как, скажем, древолазов, но сильнее, чем у всех наших жаб. Предупреждает о своей несъедобности жерлянка так: переворачивается черно – красным брюхом вверх. Когда напугана, словно пеной, покрывается ядовитыми выделениями.

 Синяя стрекоза
- это насекомые с двумя парами сильных перепончатых крыльев, с помощью которых они могут летать с большой скоростью. Часто крылья окрашены в
Синяя стрекоза
- это насекомые с двумя парами сильных перепончатых крыльев, с помощью которых они могут летать с большой скоростью. Часто крылья окрашены в  Большое коромысло
- одна из крупных стрекоз; длина ее брюшка достигает 49- 60 мм, крыльев- 45- 49мм. Размах крыльев до 10 см. Крылья желтоватые, прозрачные, с бурыми жилками; бока рыжевато – бурые с желтыми полосами. Она живет недалеко от водоёма, но нередко ее можно встретить на опушке леса или на лугу, расположенный не значительно расстояние от водоема. Большое коромысло активна с середины лета до поздней осени , предпочитает солнечные жаркие дни.
Большое коромысло
- одна из крупных стрекоз; длина ее брюшка достигает 49- 60 мм, крыльев- 45- 49мм. Размах крыльев до 10 см. Крылья желтоватые, прозрачные, с бурыми жилками; бока рыжевато – бурые с желтыми полосами. Она живет недалеко от водоёма, но нередко ее можно встретить на опушке леса или на лугу, расположенный не значительно расстояние от водоема. Большое коромысло активна с середины лета до поздней осени , предпочитает солнечные жаркие дни.


